
Seroprevalence of Bartonella henselae and ldentification of Risk Factors in China*
2020-02-29 05:47:20SONGXiuPingZHANGHaiBinLIUQiYongSUNJiMinXULeiGUShaoHuaSUNWanWanYUEYuJuanRENDongShengWANGJunandLIDongMei
SONG Xiu Ping, ZHANG Hai Bin, LIU Qi Yong, SUN Ji Min, XU Lei, GU Shao Hua,SUN Wan Wan, YUE Yu Juan, REN Dong Sheng, WANG Jun, and LI Dong Mei,#
1. State Key Laboratory of Infectious Disease Prevention and Control, Collaborative Innovation Center for Diagnosis and Treatment of Infectious Diseases, National Institute for Communicable Disease Control and Prevention, Chinese Center for Disease Control and Prevention, Beijing 102206, China; 2. The Qinghuangdao Municipal Center for Disease Prevention and Control, Qinhuangdao 066000, Hebei, China; 3. Zhejiang Provincial Center for Disease Control and Prevention,Hangzhou 310051, Zhejiang, China; 4. Ningbo Center for Disease Control and Prevention, Ningbo 315010, Zhejiang, China
Serum samples were tested for Bartonella henselae IgG antibodies using indirect immunofluorescence assays. We then analyzed associated risk factors. Serum samples were considered positive when reactive at a dilution of more than 1:320. Differences between groups and risk factors associated with Bartonella exposure were statistically analyzed using Chi-square tests and the generalized linear model. 122 of 1,260 samples(9.68%) were positive for B. henselae infection. The infection rate ranged from 0% to 30.43% and differed significantly among age groups (P < 0.01);infection rate in the 50–59 years group was significantly higher than that in other age groups.The seroprevalence of Bartonella varied significantly among sites within the four provinces, and the infection rate of field workers was significantly higher than that of urban workers.
Key words:Bartonella henselae; Serological analysis; ?ndirect immunofluorescence assay
Bartonella henselae (B. henselae) is a fastidious,slow-growing, gram-negative, hemotropic bacterium that infects blood erythrocytes and endothelial cells in its host[1]. B. henselae is responsible for cat scratch disease (CSD), bacillary angiomatosis, bacteremia, endocarditis, and peliosis hepatitis in patients with acquired immunodeficiency syndrome. Importantly, B.henselae is transmitted to humans via trauma caused by cats (cat scratches or bites) or via bites from infected fleas. Laboratory methods to diagnose Bartonella infections include culturebased isolation, serological assays,histopathological examinations, and molecular detection of Bartonella DNA in affected tissues.However, Bartonella spp. are not easily detected by routine bacterial culture protocols. Therefore, in order to determine the incidence of Bartonella infection, it is necessary to investigate the seroprevalence of this bacterium in the general population and determine the primary reservoirs and vectors involved in transmission of infection. In recent years, serological tests for detection of antibodies for different Bartonella species have been developed, and serologic testing for antibodies targeting B. henselae has been proposed as an alternative to skin testing[2].
The seroprevalence of B. henselae varies among different countries and geographic regions within countries The seropositivity rates among the healthy adults are 30% in Germany, 19.8% in Greece.Furthermore, similar or lower seroprevalence rates have been found in healthy populations from the Netherlands (3.3%)[3]. However, the seroprevalence of B. henselae in China and the risk factors for infection have not been clarified.
Accordingly, in this study, we evaluated 1,260 blood samples from healthy populations in China in order to detect anti-B. henselae antibodies using indirect immunofluorescence assays (IFAs) and then analysed the risk factors associated with infection.
Serum samples (n = 1,260) were collected from healthy populations from four regions in China(Table 1). All participants provided clinical and demographic information, including age, sex, pet contact, bite/scratch history, and living and working environments. Of the donors, 51.03% were male,and 48.97% were female (age range: 1–83 years).Serum samples were stored at ?20 °C before testing.
The experimental research reported in this study was approved by the ethics committee of National Institute for Communicable Disease Control and Prevention, Chinese Center for Disease Control and Prevention. Human research was carried out in compliance with the Helsinki Declaration. All participants provided informed consent for participation in this study.
A commercially available B. henselae-based IFA test kit (Euroimmun, Order No. FI219b-1005G,Hangzhou) was used to assess the presence of IgG antibodies against B. henselae in serum samples.Sera were diluted to 1:100, 1:320, or 1:1,000 in phosphate-buffered saline (PBS-Tween buffer;provided in the test kit), and IFAs were then conducted according to the manufacturer’s protocol.Positive and negative sera (provided in the kit) were used for quality control in each test.Immunofluorescence was observed using anepifluorescence microscope at a magnification of 400×. According to the manufacturer’s guidelines, a titer of 1:320 was considered indicative of an infection.
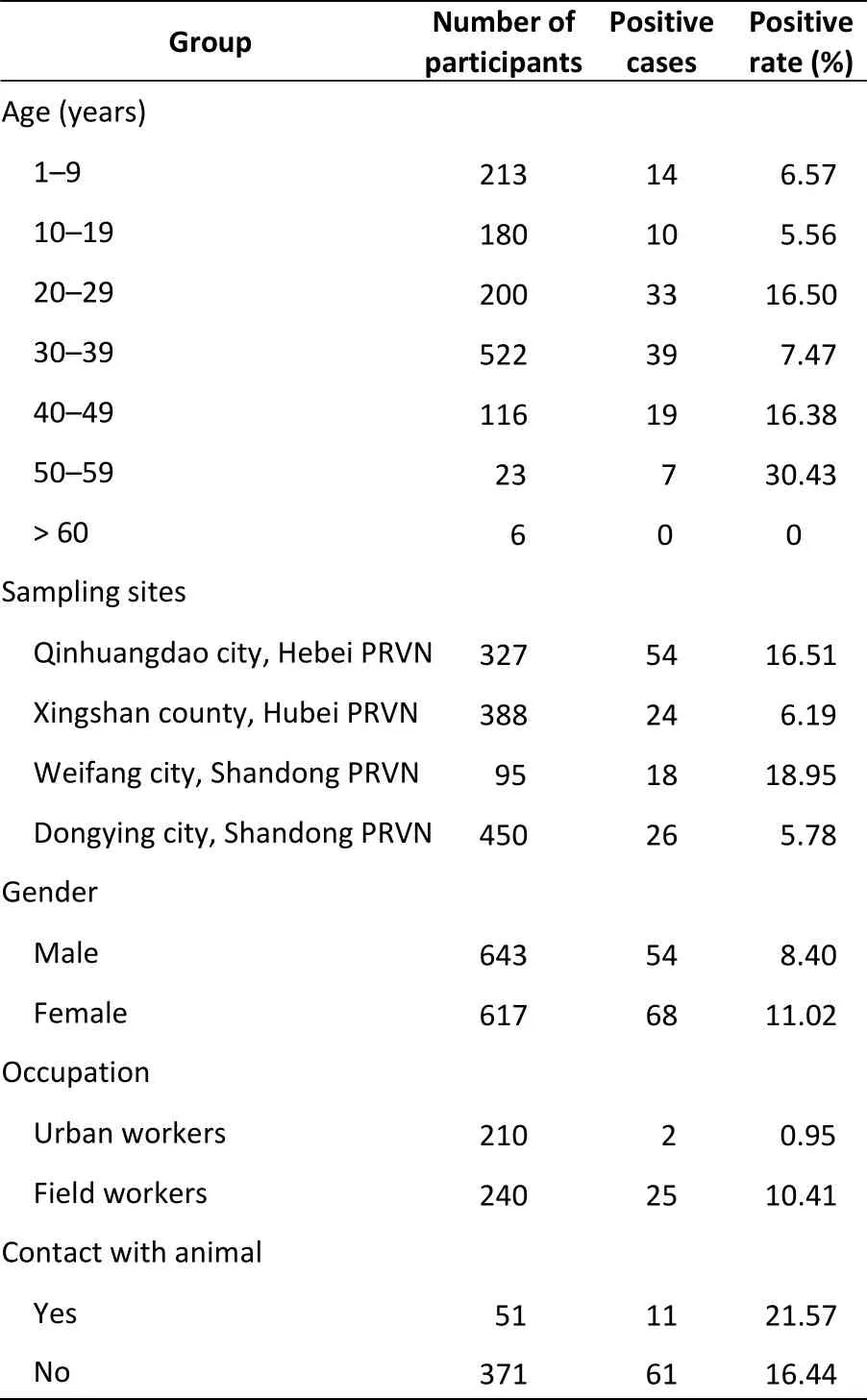
Table 1. Characteristics of serosurvey participants,in China, 2008–2013
Data were analyzed using R3.4.3[4]. Univariate analyses (Chi-square tests) were performed to determine the effects of different independent risk factors. We conducted generalized linear model(GLM) analysis with a binomial family using human prevalence data, with the following models:
where Piindicates the prevalence of B. henselae at location I; Gi, Ai, and Jiindicate the sex, age, and occupation of the patients, respectively; Liindicates the four different locations; Ciindicates whether the patient had contact with a potentially infected animal host; a indicates the overall intercept; and bnindicates the partial regression coefficient of the model.
B. henselae antibodies were evaluated in 1,260 serum samples, and the positive rate was 9.68%(Table 1). According to the results of Chi-square tests, the seroprevalence of B. henselae varied significantly among sites in the four provinces(5.78%–18.95%; χ2= 40.05, P < 0.001). The seropositivity rates in Dongying city and Xingshan county were significantly lower than those in other cities (Table 1).
Among seropositive individuals, males (8.40%)had a lower seroprevalence than females (11.02%);however, this difference was not statistically significant (χ2= 0.755, P = 0.385). Similar results were observed for the GLM (Table 1).
In our study, the highest seropositivity was observed in patients 50–59 years old (χ2= 41.22, P <0.001), and similar results were observed for the GLM. The seropositivity rates among individuals with a history of raising and being scratched by cats/dogs was not significantly different (χ2= 0.51, P = 0.48),similar to the results in the GLM (P > 0.05). The seroprevaleces of urban workers and field workers were significantly different (χ2= 16.15, P < 0.001, OR= 12.09) in the GLM (P < 0.01, Table 1).
Modern diagnostic approaches, including serological tests and polymerase chain reaction,provide precise information concerning B. henselae infection. Serological testing is the reference test for the diagnosis of CSD and other Bartonella-based infections. Although determination of the seroprevalence of B. henselae in the general population is important for controlling and preventing CSD, few large-scale epidemiological surveys of B. henselae have been reported among such populations.
To the best of our knowledge, this study report the prevalence of anti-Bartonella IgG antibodies in a large-scale population in China advancedly. We found that 9.68% of healthy populations had IgG antibodies against B. henselae; this rate was slightly lower than those reported by other investigators in other countries[3]. Some previous studies have indicated that B. henselae may be endemic in China and that its seroprevalence may be 5.78%–19.60% in Zhejiang[5]and 14.28% in Yunnan[6]. Indeed, the seroprevalence of Bartonella infection varies according to geographical region.Warm and humid environments are associated with higher seroprevalence rates for B. henselae because these areas have the highest number of potential vectors[7]. Zhejiang and Yunnan are located in the central and southwestern part of China, and the mild, wet climate is suitable for propagation of arthropod vectors. These B.henselae seropositivity rates were higher than that in our study, indicating that the climate may just be one of several factors influencing the prevalence of Bartonellosis. Therefore, new surveys should be conducted with larger numbers of participants and covering broader regions, particularly southern China.
Because of the inherent serological crossreactivity among Bartonella species in IFAs[5],the observed seropositivity for B. henselae antigen may represent a prior or current infection with another Bartonella species. According to our analysis, crossreactivity with other Bartonella species cannot be ruled out; however, the kit used in this study has more than 84% specificity and 88%sensitivity for B. henselae IgG. Thus, our data suggested that the antibodies detected in this study were representative of all Bartonella species,although most were against B. henselae. Despite this assumption, the observed 9.95% seroprevalence suggested that the rate of Bartonella infection in the general population may be underestimated in China.The epidemiology of and risk factors for Bartonella infections have not been fully elucidated. In this study, there were no significant differences in sex or animal contact, and only residential area was significantly between urban workers and field workers. Working conditions with a high probability of close contact with rodents, bacteremic cats, fleas,lice, and other potential vectors; repeated parenteral exposures; and insufficient medical care may lead to increased infection rates[5,8].Accordingly, it is recommended that field workers take appropriate protective measures to reduce bites by hosts and blood-sucking vectors. Previously published studies have indicated that B. henselae infections are more common in children than adults[9]. However, we did not observe a significantly higher seropositivity among children than adults in our study. In contrast, in our study, the highest seropositivity was observed in patients 50–59 years of age. Similar results have been reported in previous literatures[5]. With the growth of age,people has been exposed to risk factors such as cats and ticks for a longer time, and his/her immunity begins to decline, so the positive rate of infection is higher. There are few people over the age of 60 years old in this experiment, so the results are biased. In the future, we will make further investigation on the people over the age of 50 years old. Moreover, sex has been shown to not affect the prevalence of B. henselae infection[5]. Similarly, in our current study, we found no significant relationship between seroprevalence of B. henselae and sex. However, in one previous study, males were found to be more likely to acquire CSD than females[10].
B. henselae infection is a zoonotic, vector-borne disease, and animals and their parasitic vectors play important roles in the transmission of this bacterium. Cats and cat fleas are the most important hosts and vectors, respectively, although other animals and insects, including dogs, rodents, lice,and flies, play important roles as well. In our study,we found that the seroprevalence rate of healthy individuals with a pet contact history was 21.57%,which was higher than that of donors without pet contact history (16.44%); however, this difference was not statistically significant. In future studies, we will separate people based on the contact history of cats and dogs then analyze, and the results could be more accurate.
In this study, we found that Bartonella infections were widespread among healthy individuals,indicating that a substantial portion of healthy individuals may be asymptomatic carriers of B.henselae or have been infected previously with B.henselae. Because Bartonella spp. are emerging and re-emerging pathogens that can cause various symptoms, it is important to identify certain potential risk factors and dissect the degree of seropositivity to various Bartonella agents. Our current findings therefore provide an epidemiological and serological framework for future Bartonella studies.
&These authors contributed equally to this work.
#Correspondence should be addressed to LI Dong Mei,E-mail: lidongmei@icdc.cn
Biographical notes of the first authors: SONG Xiu Ping,female, born in 1972, Associate Senior Technologist,majoring in infectious disease; ZHAND Hai Bin, female,born in 1973, Associate Senior Technologist, majoring in public health research.
Received: July 23, 2019;
Accepted: December 3, 2019
 Biomedical and Environmental Sciences2020年1期
Biomedical and Environmental Sciences2020年1期
- Biomedical and Environmental Sciences的其它文章
- Contamination of Prohibited Substances in Various Food Products in Guangzhou, China*
- Strong Correlation of Abnormal Serum and Urinary lodine Levels with Papillary Thyroid Cancer:A Case-control Study*
- Optimization of Gas Chromatography-electron lonization-tandem Mass Spectrometry for Determining Toxic Non-ortho Polychlorinated Biphenyls in Breast Milk*
- Characteristics of Hypertension Death in Low-income Regions of lnner Mongolia, China*
- Analysis of Cardiovascular Risk Factors in Newly Defined Stage 1 Hypertension among Chinese on the Basis of the 2017 ACC/AHA Hypertension Guidelines*
- Effects of lncretin-based Therapies on Weight-related lndicators among Patients with Type 2 Diabetes: ANetwork Meta-analysis*