
A potential germ cell-speci fic marker in Japanese flounder,Paralichthys olivaceus: identi fication and characterization of lymphocyte antigen 75 (Ly75/CD205)*
2018-08-02 02:51:08YANGYang楊陽LIUQinghua劉清華MADaoyuan馬道遠(yuǎn)SONGZongcheng宋宗誠LIJun李軍
YANG Yang (楊陽) , LIU Qinghua (劉清華) , MA Daoyuan (馬道遠(yuǎn)) SONG Zongcheng (宋宗誠) , LI Jun (李軍)
1 Key Laboratory of Experimental Marine Biology, Institute of Oceanology, Chinese Academy of Sciences, Qingdao 266071,China
2 Laboratory for Marine Biology and Biotechnology, Qingdao National Laboratory for Marine Science and Technology, Qingdao 266071, China
3 University of Chinese Academy of Sciences, Beijing 100049, China
4 Weihai Shenghang Aquatic Product Science and Technology Co. Ltd., Weihai 264200, China
Abstract Some germ cell marker genes, such as vasa, nanos, and dead end ( dnd), have been identi fied in fish. Recently, lymphocyte antigen 75 (Ly75/CD205) has been identi fied as a mitotic germ cell-speci fic cell-surface marker in several fish species. In this study, the Japanese flounder Paralichthys olivaceus ly75 homolog ( ly75) was cloned and its expression pattern in gonads was analyzed. The full-length cDNA of ly75 was 7 346 bp, with an open reading frame (ORF) of 5 229 bp. The ORF encoded a protein containing 1 742 amino acids with a predicted molecular mass of 196.89 kDa. In adult tissues, ly75 transcripts were detected in all analyzed tissues but abundantly in the testis. In in-situ hybridization analyses, ly75 mRNA was predominantly localized in oocytes in the ovary and spermatogonia in the testis, but ly75 mRNA was not detected in oogonia, spermatocytes, spermatids, or spermatozoa. These results indicated that ly75 could be a potential germ cell-speci fic marker in P. olivaceus, as in other fishes.
Keyword: Ly75/CD205; Japanese flounder; Paralichthys olivaceus; gonads; germ cell-speci fic marker
1 INTRODUCTION
Germ cells are highly specialized cells that play an extremely important role in fish reproduction(Kurokawa et al., 2007). In teleost fish, mature gametes are acquired from primordial germ cells(PGCs) through a series of basic biological processes.Among them, the PGCs, the diploid oogonia, and spermatogonia are known as special stem cells that provide the foundation for gametogenesis through self-renewal and differentiation (Schulz et al., 2010;Nakamura et al., 2011; Lacerda et al., 2012; Kagawa,2013). However, the molecular and cellular mechanisms underlying the continuous production of mature gametes in adult fish are poorly understand.Therefore, for the molecular, cellular, and functional characterization of germ cells, germ cell marker genes such as vasa, nanos, and dead end ( dnd) have been identi fied in recent decades (Nagasawa et al., 2010; Li et al., 2015; Wang et al., 2015b). Among them, Nanos 2, a type of nanos, has been extensively studied and is regarded as a speci fic marker of germline stem cells in fish (Suzuki et al., 2007). Lymphocyte antigen 75( ly75, also called CD205) was first identi fied as a germ cell surface marker in rainbow trout (Nagasawa et al., 2010). Subsequently, ly7 5homologs have been cloned from several other fish species (Nagasawa et al., 2012, 2013; Presslauer et al., 2014). In mammals,Ly75 plays a role in the immune system and functions as an antigen-uptake receptor in dendritic cells (Jiang et al., 1995; East and Isacke, 2002). Although the role of Ly75 in fish germ cells is unknown, its amino acid sequence has a highly conserved region, and its expression is restricted speci fically to spermatogonia(SG). Therefore, it is considered to be a practical mitotic germ cell-speci fic cell-surface marker and it has been used in SG transplantation in fish (Nagasawa et al., 2010, 2012).
Japanese flounder ( Paralichthys olivaceus), a cold water benthic flat fish, is one of the most important aquaculture species in China, Korea, and Japan(Radonic and Macchi, 2009; Si et al., 2016). Due to its high commercial value, it has been intensively studied, and most research on its reproductive biology has focused on gametogenesis (Fan et al., 2014; Wang et al., 2015a). Although some germ cell markers have been identi fied in P. olivaceus, there is still much to learn about its germline cells, including their molecular and cellular makeup and functions.Therefore, in this study, we identi fied a potential germ cell-speci fic marker, lymphocyte antigen 75 ( ly75), in P. olivaceus, and investigated its gene expression pattern using real-time quantitative PCR (RT-PCR)and in-situ hybridization (ISH) analyses.
2 MATERIAL AND METHOD
2.1 Experimental animals
Nine Japanese flounders (three 8-month-old males,three adult males, and three adult females) were obtained from Oriental Ocean Sci-Tech Co., Ltd.(Shandong Province, China). The samples were collected and excised after being anesthetized with a 0.05% (w/v) solution of ethyl 3-aminobenzoate methane sulfonate (Sigma-Aldrich, Shanghai, China).To explore the function of ly75 in subsequent transplantation in marine fish, tissues including both types of testes, adult ovaries, and the gill, intestine,liver, kidney, spleen, stomach, heart, and muscle from 8-month-old males were rapidly excised and frozen in liquid nitrogen. Half of each testis and ovary sample was fixed in Bouin’s fluid and 4% paraformaldehyde in Sorensen’s phosphate buffer (0.1 mol/L, pH 7.2)for at least 24 h and then preserved in 70% ethanol.All animal work was conducted according to relevant national and international guidelines and was approved by the Institute of Oceanology, Chinese Academy of Sciences.
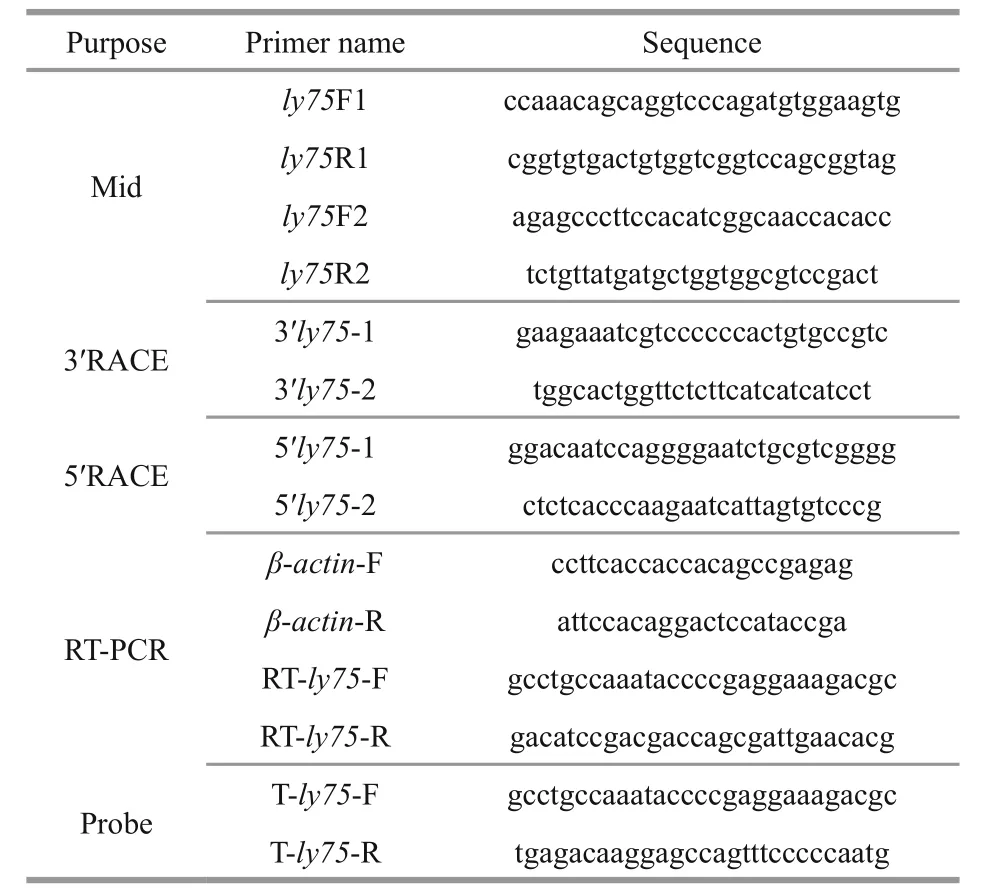
Table 1 Primers used in this study
2.2 Cloning of full-length ly75 cDNA
Total RNA was extracted from the testes of 8-month-old Japanese flounder using an RNA fast 200 kit (Fastagen Biotech, Shanghai, China). Firststrand cDNA was synthesized using a Transcript Firststrand cDNA synthesis kit (TransStart, TransGen,Beijing, China), according to the manufacturer’s instructions. The cDNA fragment of the ly75 gene was ampli fied by PCR with primers (Table 1) designed according to the highly conserved regions of ly75/ cd205 homologs from other fish species (Table 1). The PCRs were performed using KOD-Plus-Neo(Toyobo, Tokyo, Japan) in a PTC-100 thermal cycler(Bio-Rad, Hercules, CA, USA). Subsequently, 5′- and 3′-rapid ampli fication of cDNA ends (RACE) were carried out using a SMARTer RACE cDNA Ampli fication Kit (Clontech, Mountain View, CA,USA) using the gene-speci fic primers shown in Table 1. The reaction mixture contained 2 μL 10×buffer,2 μL 2 mmol/L dNTPs, 1.4 μL 25 mmol/L MgSO4,0.6 μL forward and reverse primers (10 μmol/L),11 μL ddH2O, 0.4 μL KOD-Plus-Neo (1 U/μL), and 2 μL cDNA. The thermal cycling conditions were as follows: 2 min at 94°C; 10 s at 98°C, 30 s at T m °C,and 1 min at 68°C for 40 cycles; and finally 10 min at 72°C. The molecular mass and p I of predicted poly75 polypeptide were estimated using the compute pI/Mw tool (http://web.expasy.org/compute_pi/).
2.3 Phylogenetic and bioinformatics analyses
Homology searches of the nucleotide and deduced amino acid sequences of ly75 were conducted at the National Center of Biotechnology Information website (http://www.ncbi.nlm.nih.gov/). The deduced amino acid sequences were aligned using AlignX in Vector NTI 11.5 Suite (Life Technologies, Carlsbad,CA, USA). The percentages of similarity and identity among fish Ly75 homologs were calculated using LALIGN (http://www.ch.embnet.org/software/LALIGN-form.html). A phylogenetic tree was constructed using Mega 4.1 software with bootstrap analysis of 1 000 replicates, by the neighbor-joining method. The signal peptide sequences were determined by SignalP (http://www.cbs.dtu.dk/services/SignalP/). A domain structure analysis was carried out using SMART (http://smart.emblheidelberg.de/). Transmembrane helices in proteins were predicted by TMHMM (http://www.cbs.dtu.dk/services/TMHMM/).
2.4 Semi-quantitative RT-PCR analysis
Total RNA was extracted from tissue samples(including testis, ovary, gill, intestine, liver, kidney,spleen, stomach, heart and muscle) as described above. Speci fic primers (Table 1) were used to amplify ly75 and β- actin by RT-PCR, and the primer speci ficity for each gene was veri fied by sequencing. The cDNA was synthesized from 1 μg total RNA using the PrimeScript RT reagent kit (TaKaRa, Dalian, China)following the manufacturer’s instructions. The reaction mixture contained 12.5 μL 2×buffer, 9.5 μL ddH2O, 0.5 μL forward and reverse primers(10 μmol/L), and 2 μL cDNA. All samples were run in triplicate. The thermal cycling conditions were as follows: 3 min at 95°C; 5 s at 95°C and 30 s at 62°C for 35 cycles; and 30 s at 95°C, 20 s at 60°C; and 30 s at 72°C, followed by a final elongation step at 72°C for 3 min. The PCR products were electrophoresed on 2% (w/v) agarose gels.
2.5 Histology
The testes and ovaries fixed in Bouin’s fluid were dehydrated with an ethanol gradient, embedded in paraffin, and cut into a series of sagittal and cross sections (5 μm thick). The tissue sections were stained with hematoxylin and eosin (HE) for histological observations.
2.6 In-situ hybridization
The localization of ly75 transcripts was analyzed by section in-situ hybridization (SISH), as described by Wang et al. (2015b). Samples were dehydrated using a methanol gradient, embedded in paraffin wax,and cut into 8-μm thick sections. Probes for ly75(Table 1) were individually synthesized using a DIG RNA Labeling Kit (SP6/T7) (Roche, Mannheim,Germany) following the manufacturer’s instructions.
2.7 Immunohistochemistry
The localization of Nanos 2 was analyzed using immunohistochemical analyses. Samples were dehydrated with a methanol gradient, embedded in paraffin wax, and cut into 8-μm thick sections. The tissue sections were treated with lead citrate (BBI,Shanghai, China) for antigen retrieval and 3% H2O2to block endogenous peroxidase. The primary antibody against Nanos 2 protein (BBI, China) was diluted 1:200 and incubated with the samples at 4°C overnight. The secondary antibody was goat antirabbit IgG (ABclone, China), which was diluted 1:800 and incubated with the samples for 30 min at room temperature.
3 RESULT
3.1 Cloning of ly75 from Japanese flounder
The full-length cDNA of ly75 (GenBank accession No. MF983803) was 7 346 bp, with an open reading frame (ORF) of 5 229 bp. The ORF encoded a protein consisting of 1 742 amino acids with a predicted molecular mass of 196.89 kDa. The deduced amino acid sequence of ly75 showed 70.7% identity and 90.1% similarity with that of Cynoglossus semilaevis,49.3% identity and 74.9% similarity with that of zebra fish, 61.4% identity and 83.9% similarity with that of medaka, and 35.8% identity and 67.1%similarity with that of mouse. A phylogenetic tree analysis (Fig.1) revealed that P. olivaceus Ly75 was associated with teleost Ly75s and belonged to the Ly75 protein subfamily. In addition, analyses of the protein sequence of ly75 (Fig.2) revealed a predicted N-terminal signal peptide cleavage site between amino acid positions 28 and 29. And the sequence contained a cysteine-rich domain (RICIN), a fibronectin type II (FN2), 10 C-type lectin-like domains (CLECT), and a transmembrane domain (TM).
3.2 Analyses of ly75 mRNA transcript levels in different tissues by RT-PCR
Transcripts of ly 75 were abundantly detected in the testis of 8-month-old fish, and in the gill, kidney,spleen, stomach, and heart. Transcripts of ly 75 were detected at low levels in the adult testis and ovary,intestine, liver, and muscle (Fig.3).

Fig.1 Phylogenetic tree of deduced amino acid sequences for Ly75 from Japanese flounder and other vertebrates constructed by MEGA 4.1 using neighbor-joining method
3.3 Detection of ly75 mRNA in gonads by ISH analysis
The mRNA of ly75 was detected in the testis and ovary by ISH. The localization of the Nanos 2 protein was also determined using immunohistochemical analyses. In adult ovaries, ly75 mRNA was predominantly located in the cytoplasm of oocytes at stage III and IV, and could not be detected in oogonia and oocytes at stage II (Fig.4a, b). In the testis, ly75 mRNA was predominantly detected in spermatogonia,and could not be detected in spermatocytes,spermatids, or spermatozoa (Fig.4f, g). The Nanos 2 protein was expressed in germline stem cells:spermatogonia in the testis (Fig.4j) and oogonia in the ovary (Fig.4d).
4 DISCUSSION
In this study, we isolated and characterized a ly75 homolog from P. olivaceus. The deduced amino acid sequence of ly75 was similar to that of ly75s of other teleosts, showing 90.1% similarity to that of C. semilaevis, 74.9% similarity to that of zebra fish(Presslauer et al., 2014) and 67.1% similarity to that of mouse (Jiang et al., 1995). The Ly75 amino acid sequence contained typical conserved domain structures including one RICIN, one FN2, 10 CLECT,and one TM, with high identities to those of other Ly75 proteins (Nagasawa et al., 2012). The phylogenetic tree constructed using the neighborjoining method showed that the Ly75 protein was closely related to the Ly75 clade. Overall, these results revealed that the clone identi fied from P. olivaceus was a Ly75 homolog.

Fig.2 Molecular characterization of Japanese flounder ly75
The RT-PCR analyses of ly75 transcript levels in different tissues revealed that ly 75 mRNA was present in the testis, gill, and heart. Similar results were also reported for rainbow trout (Nagasawa et al., 2010),paci fic blue fin tuna (Nagasawa et al., 2012), and Atlantic salmon (Nagasawa et al., 2013). This wide expression pattern was possibly related to the function of this protein in the immune system, as previously found in mammalian species (Jiang et al., 1995).Similar to the case in rainbow trout (Nagasawa et al.,2010) and Blue fin tuna (Nagasawa et al., 2012), ly75 transcripts were abundant in the testis at the early development stage. The strong expression in the gonads indicated that ly75 may play a key role in early gametogenesis in P. olivaceus.
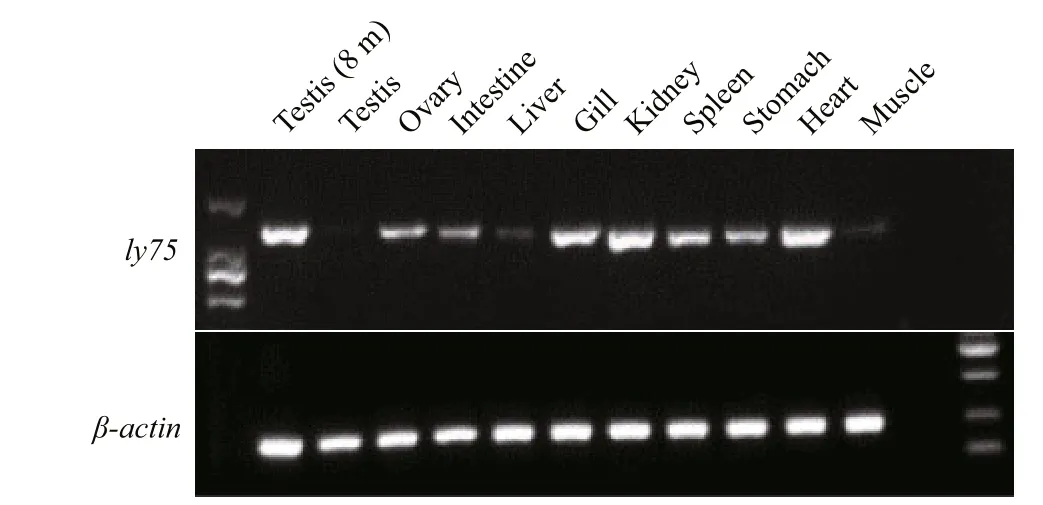
Fig.3 Tissue-speci fic expression of ly75 mRNA as determined by RT-PCR (internal control, β- actin)
The ISH analyses showed that ly75 mRNA was present in oocytes at stage III and IV, but not in oogonia and oocytes at stage II and V in ovaries. In contrast, ly75 transcripts were detected in oogonia and nucleolus-stage oocytes in rainbow trout(Nagasawa et al., 2010), and in oocytes at stages I, II,and III in zebra fish (Presslauer et al., 2014). Most fishes release eggs, and fish embryos rely upon the maternal provision of immunity molecules for protection against invading pathogens before their own immune system fully develops (Zhang et al.,2013). Therefore, in oogenesis, maternally transferred immunity molecules including lysozyme, lgM, and the egg yolk protein phosvitin are accumulated and stored in oocytes (Wang et al., 2017). Based on the present results, Ly75 as an immunity-related protein expressed in oocytes, may be related to maternally derived immunity molecules. In the adult testis, ly75 mRNA was predominantly detected in SG, just its homologs were only detected in type A SG in rainbow trout (Nagasawa et al., 2010), zebra fish (Presslauer et al., 2014), and Thunnus orientalis (Nagasawa et al.,2012).
Immunohistochemical analyses revealed that the Nanos 2 protein was predominantly expressed in germline stem cells: spermatogonia in the testis and oogonia in the ovary, as in other fish (Suzuki et al.,2007; Huang et al., 2017). Several other genes have been reported as germ cell-speci fic markers in fish. In our previous study, dnd was identi fied as a gene essential for PGCs migration and oocytes and spermatocytes (Wang et al., 2015b). The results of the present study indicate that ly75 could be a potential germ cell-speci fic marker in P. olivaceus,since it is speci fically expressed in germ cells. In future research, we intend to functionally characterize this protein.
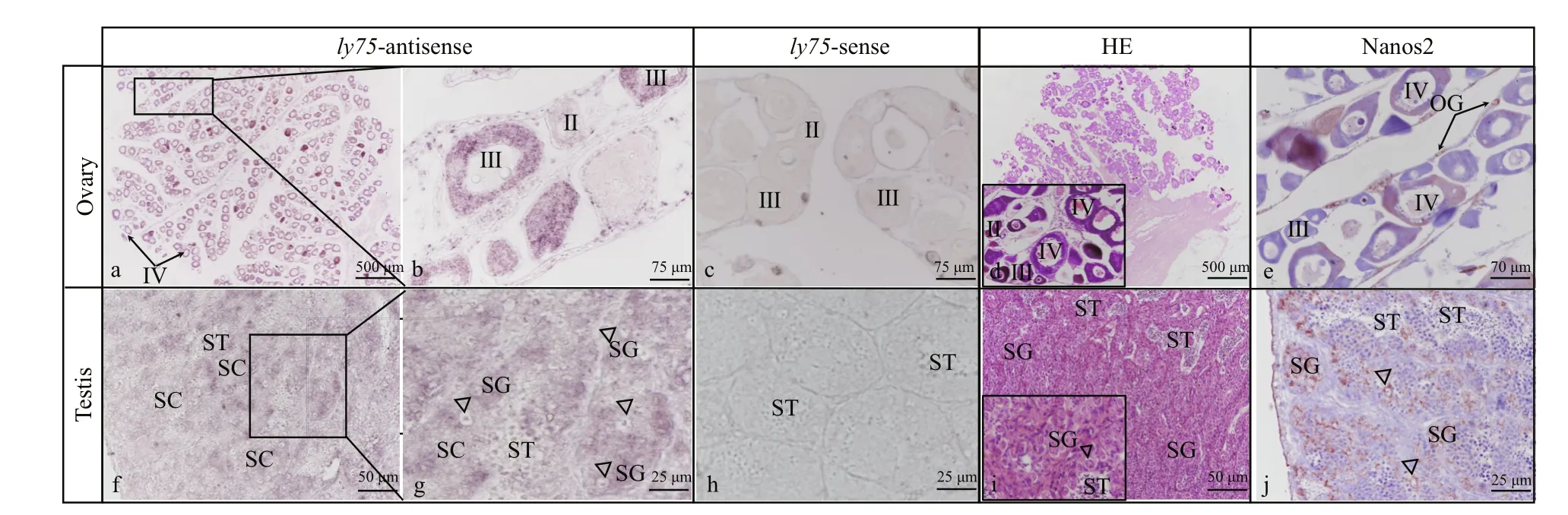
Fig.4 Distribution of ly75 transcripts in adult P. olivaceus testis and ovary
5 CONCLUSION
We cloned Japanese flounder ly75 and analyzed its expression pattern. In RT-PCR analyses, ly75 transcripts were detected in all analyzed tissues but abundantly in the testis. The ISH analyses showed that ly75 mRNA was predominantly localized in oocytes in the ovary and spermatogonia in the testis. We did not detect ly75 mRNAs in the oogonia, spermatids,spermatocytes, or spermatozoa. Therefore, our results indicate that ly75, like dnd, could be a potential germ cell-speci fic marker in P. olivaceus.
7 DATA AVAILABILITY STATEMENT
The datasets generated and analyzed during the current study are not publicly available from NCBI(https://www.ncbi.nlm.nih.gov/) until Sep. 3, 2018 due to author request, but are available from the corresponding author on reasonable request.
 Journal of Oceanology and Limnology2018年4期
Journal of Oceanology and Limnology2018年4期
- Journal of Oceanology and Limnology的其它文章
- Editorial Statement
- Effects of seawater acidi fication on the early development of sea urchin Glyptocidaris crenularis*
- Dietary effects of A zolla pinnata combined with exogenous digestive enzyme (Digestin?) on growth and nutrients utilization of freshwater prawn, Macrobrachium rosenbergii(de Man 1879)
- Preliminarily study on the maximum handling size, prey size and species selectivity of growth hormone transgenic and non-transgenic common carp Cyprinus carpio when foraging on gastropods*
- Hydrodynamic characteristics of the double-winged otter board in the deep waters of the Mauritanian Sea*
- De novo transcriptome sequencing reveals candidate genes involved in orange shell coloration of bay scallop Argopecten irradians*